
Sex Determination
Contents of Table
- Abstract
- Introduction
- Historical Development
- What is sex Determination
- What is Fertilization
- What is Chromosomes
- Sex Determination with Chromosomal Machanisms
- Sex determination Systems
- Conclusion
Abstract
A wide array of sex determination mechanisms, encompassing genetic and non-genetic pathways (i.e., hormonal, environmental, and epigenetic factors), have been found among different organisms. The presence of two complementary sexes, male and female, is an ancient feature in biology. Triggering the differentiation of male and female reproductive organs is a conserved ontogeny process, and sex determination is an inherently fascinating process. Sex determination is dependent on molecular signaling whether the male and the female differentiating pathway is activated, and different triggering elements such as genetic, non-genetic, and epigenetic factors control the whole process. This chapter describes various aspects of sex determination, such as historical development, the evolution of sex chromosomes, and different sex determination systems in other organisms.
Sex determination, although crucially important for the survival of a species, is arguably the least conserved mechanism in evolution and is instead one of the most variable developmental pathways. There are two main mechanisms of sex determination: genetic (GSD) and environmental sex determination (ESD). Among st vertebrates, many reptiles such as crocodiles and turtles determine sex by ESD, whereas birds and mammals, including human display GSD. GSD can be further subdivided into those for which the males are hetero gametic, that is, an XX–XY system, and those with heterogametic females, that is, ZZ–ZW system. Here, we review genetic mechanisms of sex determination.
Introduction
Sexual reproduction is a historical process of life on earth, and the most popular heterogametic system (X and Y sex chromosomes) in humans and many other organisms leaves an imprint that sex determination mechanism is ancient and conserved . Sex determination is inherently an integral part of reproduction that separates reproductive organs responsible for male and female gamete production . It is an intricate developmental process that describes whether the individual will be developed as male or female. At the same time, sexual differentiation is the subsequent development of phenotypic differences (primary and secondary sexual characters) between male and female individuals from an undifferentiated zygote . Sex differentiation stages are decided by the sex determination, that is, the gender-specific response of different tissues to hormones produced by the gonads (male or female reproductive organs) distinctly in both genders .
Various pathways decide males and females, and these pathways have been evolved rapidly in many species or genera/taxa. Sex determination is regulated by several different genetic (i.e., sex chromosomes) and non-genetic pathways (i.e., hormonal, environmental, and epigenetic factors). In different animal and plant species, genetic systems have been classified as homogametic or heterogametic sex types . In most species, heteromorphic sex chromosomes are present, which are the results of evolutionary changes in size or shape of the sex chromosomes. Similarly, non-genetic pathways also play a key role in determining the sex in fern species (hormonal regulation) and crocodiles, alligators, and turtles (thermo-regulation).
Sex determination is an important evolutionary process as it encourages the genetic fitness of an individual. The ultimate aim of sex determination is to promote the heterozygosis or accumulation of diverse alleles in a species, which is vital for creating genetic variation in living organisms. It is pivotal in plant breeding to design a specific breeding program as per the need and demands of diverse stakeholders to improve plant productivity and nutritional quality. The exact mechanism or genes determining the sex or reproductive organ is unclear. Therefore, it is an important area of study in developmental and evolutionary biology, as well as in ecology. Sex determination in various plant and animal species is not under the control of the universal model. Thus, this chapter provides a brief overview of the different mechanisms of sex determination in plant and animal species.
C. elegans has two sexes, hermaphrodite and male. The hermaphrodite is a modified female that in the fourth larval stage makes and stores sperm to be used later to fertilize oocytes produced within the gonad of the same animal after spermatogenesis is finished. The embryos produced by self-fertilization are encased in an egg shell and initiate development within the uterus of the hermaphrodite. When they reach about the 30-cell stage, the egg-embryos are laid by the hermaphrodite through a vulva. A useful consequence of this mode of reproduction is that a single hermaphrodite heterozygous for a recessive gene automatically generates one-quarter recessive homozygote’s in its brood of self progeny—a feature, shared with Mendel’s peas, that helped attract Sydney Brenner to the worm in the first place. At the same time, Brenner saw that males, which can mate with and transfer their sperm to hermaphrodites to produce cross progeny, are useful to the experimentalist for making new combinations of genes. Presumably this is also why C. elegant has retained the male sex, which in the short term at least is completely dispensable for reproduction.
Hermaphrodites are normally diploid, with five pairs of autonomies and two X chromosomes. Males have the same five pairs of autonomies but only a single X chromosome. Nearly all gametes—sperm and eggs—produced by hermaphrodites are haplo-X and thus give rise to XX hermaphrodite self progeny, but rare males are generated through spontaneous X chromosome loss. Males produce equal numbers of haplo-X and nullo-X sperm, so that half of the cross progeny they sire will also be male.
Males and hermaphrodites are distinctly different creatures. The first three chapters in this section describe, at the level of individual cells, the major differences between the sexes and how they arise developmentally. The embryonic cell lineages in the two sexes are essentially identical, although a few cells in each sex are programmed to die sex-specifically during late embryogenesis; for example, males get rid of two cells that in hermaphrodites would become neurons required for egg-laying. But most differences between the sexes arise during postembryonic development through different patterns of cell lineage. Surprisingly, the different patterns of lineage and differentiation are driven largely by the same genes in the two sexes, although by different cell-specific patterns of gene expression. The same multiple transcription factors and core set of intercellular signal transduction systems are used repeatedly in the sex-specific developmental pathways of both sexes.
In the first chapter of this section, Scott Emmons describes those aspects of development that are specific to the male. Much of the interesting male somatic development occurs in the tail, which in the adult contains male-specific neurons, muscles, and epidermal cells that enable the male to copulate efficiently with hermaphrodites. For the development of these tissues, some blast cells common to the two sexes initiate male-specific cell lineages. In other cases, the same cells in the two sexes differentiate differently. The major male mating structures form in dramatic morphogenetic events just before the last larval molt. Although the male somatic gonad differs substantially in overall morphology from that of the hermaphrodite, the cell lineages that give rise to the two somatic gonads are clear variants of each other.
In his chapter, Michael Herman focuses on cell fate specifications that occur only in hermaphrodites. Both sexes make use of Hox genes and asymmetric distributions of the Wnt pathway transcription factor POP-1/Tcf for patterning their anterior-posterior body axes, but these regulators are interpreted differently in the two sexes. The M cell, for example, gives rise in hermaphrodites to (among other cell types) muscles needed in the mid-body region for egg laying, whereas the M cell in males gives rise to muscles in the tail needed for copulation.
Paul Sternberg has written about a single hermaphrodite-specific organ, the vulva, which forms during larval development and provides an opening between the uterus and the external environment. A remarkably detailed description of the molecular events and individual cells involved in this process has emerged. Although the cell lineage that gives rise to the vulva is invariant, it depends critically on three standard intercellular signaling pathways: EGF-Ras-MAP kinase, LIN-12/Notch, and Wnt.
The descriptions of the differences between males and hermaphrodites naturally lead to the question of what makes them different. David Zarkower explains in his chapter that the difference between male fate and hermaphrodite fate for somatic cells is determined cell autonomously by a single master regulator, the transcription factor TRA-1: hermaphrodite fate is specified when TRA-1 is active, and male fate is specified when TRA-1 is inactive. This leads to two further questions, which Zarkower addresses: what makes TRA-1 active in hermaphrodites and inactive in males, and what are the targets of TRA-1 action? The answer to the first question involves a cell-no autonomous, global sex determination pathway, which is fairly well understood and triggered by an assessment of the ratio of the number of X chromosomes relative to the number of autoscores, the X:A ratio. The second question presents a large gap in our understanding, since very few TRA-1 targets have so far been identified.
In their chapter, Ronald Ellis and Tim Schell point out that sex determination in the germ line is not a simple recapitulation of the regulation by TRA-1 that takes place in the soma. Although the same members of the global sex determination pathway that act in the soma are required for sex determination in the germ line, the pathway operates slightly differently, and TRA-1 is not the sole final arbiter of sexual fate. In addition, as one might expect, certain germ line-specific genes are needed to control germ cell fate.
Sex determination evolves rapidly, and Eric Haag’s chapter is based on the idea that our detailed understanding of sex determination in C. elegant makes it an attractive subject for studies in comparative biology. Only two C. elegant genes are known to be related to genes with sex-specific roles in a wide range of animals: mab-3 and mab-23 affect some male cell fates and belong to the DM domain transcription factor family, along with the Drosophila gene doubles and some vertebrate genes that also act sex-specifically. Because C. elegance sex determination has otherwise evolved very rapidly, the most useful inter-species comparisons are with other nematodes, as Haag indicates. Such studies should expand as the sequences of more nematode genomes become available.
No story of sex would be complete without a discussion of dosage compensation, and in the final chapter in this section, Barbara Meyer describes how hermaphrodites assemble a protein dosage compensation complex (DCC)—which is related to a chromosome condensin complex—all along their X chromosomes to dampen X gene transcription just enough to make it equal to that found in X0 males. A functional DCC is not made in males owing to the repressive effect of the gene xol-1, which is active in males and not in hermaphrodites. How xol-1 is activated only in males is an interesting story told by Meyer involving X-linked repressors and autosomal activators that enable xol-1 to respond appropriately to the X:A ratio. The status of xol-1 expression determines both the sex of the animal—by affecting tra-1 expression via the global sex determination pathway—and whether or not a DCC will be formed.
Historical Development
Whether a plant or animal will become a male, a female, or bisexual is determined during the initial development of an organism. Hundreds of years ago, researchers have started studying the mechanism of sex determination. For example, in 335 B.C.E., Aristotle anticipated that sex is controlled or ruled by the heat of the male partner during intercourse. A male child will be born when the male parent’s heat overwhelms the female parent’s coldness and vice versa. Environmental theories of sex determination (i.e., Aristotle’s theory, in reptiles, temperature during embryo development regulates the gender) were popular until about 1900 and meticulous scientific research began after the discovery of sex chromosomes during 1900, and gradual scientific improvements followed during the next century. In 1891, Hermann Henking [6], a German Biologist studying spermatogenesis in the insect firebug (Pyrrhocoris apterus; 2n = 24), detected that, as a result of meiosis, half of the spermatozoa have not received all 12 chromosomes and ended with 11 chromosomes only. It means one of the chromosomes was not involved in meiosis. This chromosome seemed and behaved differently from others; he was not able to speculate the significance of this element and named it “X element or X body”. In 1902, Clarence Erwin McClung [7] cleared the Henking assumption through cytological observations on several grasshopper species and demonstrated that the somatic cells in female grasshoppers are different in chromosome number than do corresponding cells in the male, which he referred to as “accessory or supernumerary chromosome” and demonstrated their association with sex determination [8]. Later on, American geneticist Edmund Beecher Wilson (1905) observed differences, either in the presence or in the absence of one chromosome or in the size of one chromosome pair in germ cells of both the sexes of praetor species [9]. Another American geneticist Nettie Maria Stevens (1905) studied germ cells of both sexes of mealworm beetle (Tenebrous monitor) and found that in males, one chromosome was smaller than the other chromosome, and she confirmed that this chromosome must be regulating the sex in males [8]. Later on, this smaller chromosome was named as “Y chromosome” and the larger one as “X chromosome” by Stevens (1905). After Stevens died in 1912, Wilson was the first to designate the name “sex chromosome” for the pair of XX and XY chromosomes. Hermann Joseph Muller (1914), an American geneticist, speculated that differentiation of sex chromosomes would arise from lack of recombination due to the appearance of sex-determining genes on the Y or W chromosomes [10]. By the end of the 1950s, the male-determining function was established on the small arm of the Y chromosome and was named “testis determining factor” (TDF in humans and Tdf in mice). Further, Ohno (1967) proposed the concept of ancestral sex chromosomes and their progress to evolve modern-day sex chromosomes by degeneration of the Y or W chromosomes . As science progressed, the major breakthrough in sex determination was achieved through sequencing and transgenic approaches. Gene-sequencing approaches revealed an open-reading frame (ORF) coding a single exon gene in a male mouse (XY) and named it as a sex-determining region of the Y chromosome (SRY in humans and Sry in mice). Conclusive evidence of functionality of SRY gene was developed through transgenic approach by generating sex-reversed mouse (transgenic XX mouse having Sry gene).
What is sex Determination
A sex-determination is the biological system that determines the development of sexual characteristics in an organism. There are two different types of sexes – male and female. Chromosome plays an essential role in determining the sex of an individual.
Let us have a glimpse into sex determination and who determines the gender of the baby- father or mother.
Comparatively, sexual reproduction is much more complicated than asexual reproduction. Sexual reproduction is a slow and lengthy process. The process begins with gamete formation in the male and female bodies. Through sexual intercourse, these male and female gametes meet and fuse to form a unicellular zygote. It continues with multiple divisions of the zygote to form a multicellular embryo which eventually develops into a complete fetus. The entire process takes nine months to deliver a baby.
What is Fertilization?
Fertilization is the process in which two gametes (sex cells) – sperm and ova – fuse together to form a new cell called the zygote. During this unbiased event, all genetic information from the parents to the child has been transferred. Also, the gender of the child is determined, and the father of the child is responsible for sex determination.
What are Chromosomes?
Chromosomes are a thread-like structures holding genetic information. It is present in the cell nuclei. A normal cell contains 23 pairs (46n) of chromosomes. Out of these, one pair (2n) is sex chromosomes. Sex chromosomes are denoted as X and Y. A male cell contains XY sex chromosomes, while a female cell contains XX sex chromosomes.
A gamete contains only 23 chromosomes (23n). A sperm contains either an X or Y chromosome, i.e. there will be two sets of sperms. One set of sperm consists of only X chromosomes, while another set consists of only Y chromosomes. An ovum always contains an X chromosome. The sex of the child depends on the kind of sperm that fuses with the ovum.
During sexual intercourse, the father ejects both types of sperms, i.e. X chromosome and Y chromosome sperms.
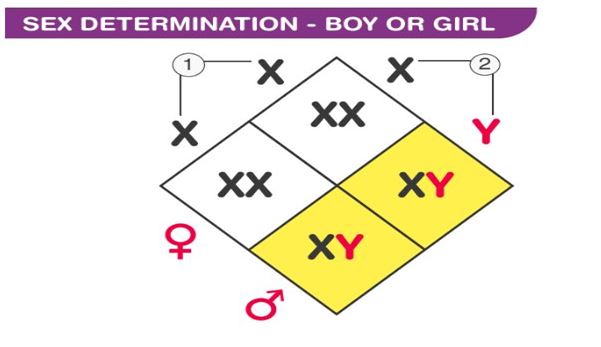
- If the sperm with an X chromosome fertilizes the ovum of the X chromosome, the zygote would have XX chromosomes, and the child will be a girl.
- If the sperm with the Y chromosome fertilizes the ovum of the X chromosome, then the zygote would have XY chromosomes, and the child will be a boy.
Sex Determination with Chromosomal Machanisms
Humans have 46 chromosomes. The total number of chromosomes are divided into 23 pairs. Out of these 23 pairs, 22 are called autoscores, whereas one pair is referred to as a sex chromosome. Males and females both possess two sets of sex chromosomes. This particular combination is useful in determining a person’s biological gender.
Male and female species differ in terms of the number or structure of their sex chromosomes (X and Y chromosomes). However, autoscores are identical in both sexes.
- All eggs would have the X chromosome as their 23rd chromosome; however, sperms might have either the X or Y chromosomes as their 23rd chromosome.
- If a sperm containing a chromosome fertilizes an egg, the resulting zygote develops into a female offspring.
- If an egg is fertilized with a sperm carrying the Y chromosome, the resulting zygote develops into a male offspring.
XX Female-XY Male Type
- Both sexes have an even number of sex chromosomes in this kind of sex determination:
- Females have two X chromosomes (XX)
- Males have one X chromosome and one Y chromosome (XY).
- Females are homogametic (XX), producing ova (female gametes) with one X chromosome.
- Males are heterogametic, producing two distinct types of sperms (male gametes). During gametogenesis 50% sperms contain X chromosome and 50% sperms contain Y chromosome.
- During fertilization, when X ova are fertilized with X sperm, XX females are produced; when X ova are fertilized with Y sperm, XY males are produced.
- This XX female/XY male sex determination process is found in humans, Drosophila, and plants.
- Various species have different chromosomal mechanisms for determining sex.
Conclusion
The several mechanisms of sex determination reveal the diverse pathways governing sex determination in both plants and animals and these pathways are also very well understood in various model organisms. The highly evolved system of sex determination is heterogametic sex determination in animals, that is, XX/XY. Even though there are several unsolved mysteries related to the sex determination system such as why heterogamete is more common in male then female? Why degeneration of sex chromosomes occurs only in few organisms not in all? With the progress in molecular techniques over the past decades, several puzzles were solved like discovery of Sry and a ray of hope arises to learn more about molecular basis of sex determination, evolution of sex chromosome, mapping of gene, sequencing, gender-dependent expression of sex-regulating gene, and relationship between the evolution of genetic degeneration and dosage compensation. Multiple “-omics” data and integrative approaches will allow scientists to address the unresolved questions and finding the new sex-determining genes as well as genetic networks involved in sex determination.
Sex refers to the difference in genetics between two members of the same species. Thus, an individual can be either male or female under normal circumstances. Scientists have revealed that definite sex determinants, also known as heterosomes, are used to transmit sex. The sex chromosomes are found in both male and female cells in the human body. A woman possesses two X-chromosomes, whereas a man has one X- and one Y-chromosome. This indicates that the sperm cell type that fertilizes the egg determines the sex of the baby. When an egg is joined with a sperm containing a Y-chromosome, a male zygote develops; when an egg is joined with sperm with an X-chromosome, a female zygote develops.
Click for further
information
https://a-zstudentpdfprojectfilesolutions.com/